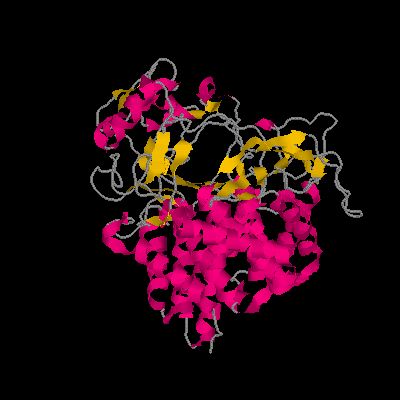
 GH48の立体構造、ちょうど中央にトンネルが見え、ここへ基質であるセルロース鎖が入る。(Clostridium cellulolyticum 由来、PDB 1FAE、RasMolを使用して作成)
GH48の立体構造、ちょうど中央にトンネルが見え、ここへ基質であるセルロース鎖が入る。(Clostridium cellulolyticum 由来、PDB 1FAE、RasMolを使用して作成)
プロセッシブ酵素とは
酵素の作用様式として、古くからエンド型とエキソ型の2つの酵素が知られています。エンド(endo)とは内側を意味し、セルラーゼの場合、セルロース鎖をランダムに切断する酵素をエンドグルカナーゼ(EC 3.2.1.4)としています。一方、エキソ(exo)とは外側を意味し、同様に、セルロース鎖を端のほうから順に切断する酵素をエキソグルカナーゼと呼んでいます。セルラーゼの場合、エキソグルカナーゼの反応生成物がセロビオースなので、セロビオヒドロラーゼと呼んでいます。(セロビオヒドロラーゼ EC 3.2.1.91の定義では、非還元末端側からセロビオース単位で切り出す酵素と記述されていますので、Trichoderma の系の主要な酵素であるファミリー7に属しているCBHIのように、還元末端側から切断する酵素については、EC 3.2.1.91には該当せず、最近になり新たに、EC 3.2.1.176が割当されました。)これらのエキソ酵素は、グルコアミラーゼのようなエキソ酵素(活性中心が穴状になっている)とは異なり、セルロース鎖が入っていく活性中心がトンネル状になり反対側に抜けていることが特徴です。グルコアミラーゼの場合、アミロースの端に酵素がついたあと、これを切断して、生じたグルコースを放出する際に、一度アミロース鎖から酵素が離れる必要があります。しかし、セロビオヒドロラーゼの場合、活性中心がトンネル状になっているので、生じたセロビオースが、トンネルの反対側から出て行くことができます。このため酵素がセルロース鎖から離れる必要がありません。このような反応様式をもつ酵素をプロセッシブ酵素(processive enzyme)と呼んでいます。
プロセッシブ酵素としては、ファミリ-の6、7、9、48の酵素が知られています。ファミリー6や7では、活性中心付近がトンネル状になっているものは、セロビオヒドロラーゼ活性を示し、活性中心付近のクレフトが、トンネル状になっていないものは、エンドグルカナーゼ活性を示します。このように、セロビオヒドロラーゼ活性とエンドグルカナーゼ活性は、セルロース鎖が結合する酵素のサブサイトと活性中心付近の構造に依存していことになります。トンネルの屋根にあたる部分は、ループから形成されており、このループのあるなしが活性の様式を分けていることになります。一方、トンネル部分の開閉がセロビオヒドロラーゼで認められていますので、セロビオヒドロラーゼは、トンネル部分が開いた状態で、エンド活性、閉じた状態でエキソ活性という二つの活性を示すと考えることができます。ファミリー6では、非還元末端側より、ファミリー7、48では還元末端側よりセロビオースを遊離することが知られています。還元糖生成量と可溶性セルロース鎖分解における粘度低下を比較しますと、エンド型の活性では、還元糖生成量よりも粘度低下が急速に起こります。一方、エキソ型の活性では、還元糖量の増加はおきますが、粘度の低下はあまりおきません。この中間にあるような酵素については、エンド-プロセッシブ(endo-processive)酵素という場合があります。
Trichoderma reeseiの系において、CBHIすなわちファミリー7のセロビオヒドロラーゼ(Cel7A)が、セルロースの分解において重要な成分であり、Clostridium thermocellumのセルロソーム系においてもCelS(Cel48A)、すなわちファミリー48(GH48)のセロビオヒドロラーゼがセルロソームの主要な成分であること、Clostridium cellulovorans 、Clostridium cellulolyticum、Clostridium josui、Clostridium acetobutylicum、のいずれのセルロソーム遺伝子クラスターにおいても、骨格タンパク質スキャフォールディン遺伝子の次は、すなわち、その下流には、GH48がコードされています。また、セルロソーム系ではない、Clostridium stercorarium、Ruminococcus albus においてもGH48を失うとセルロース分解性が低下するという報告があります。これらのことからプロセッシブ酵素のセルロースの分解における重要性がみえてくるような気がします。しかしながら、Clostridium phytofermentansではGH48の酵素がないことも知られており、かならずしもセルロースの分解にGH48が必要不可欠ではないと考えられます。
最近になり、五十嵐先生(東京大)が撮影した高速AFMの画像では[1]、CBHIは、確かにセルロース繊維上を結合したまま一方向に移動しており、活性のない変異体では移動しないことから、加水分解しながら移動するというプロセッシブな作用様式が実際に証明されています。
従来の多くの文献で、セルロースの酵素分解を紹介する文面で、エンドグルカナーゼ(EC 3.2.1.4)とセロビオヒドロラーゼ(EC 3.2.1.91)の相乗作用という表現がされていますが、これからは、セロビオヒドロラーゼは、還元末端から切断するセロビオヒドロラーゼ(EC 3.2.1.91)と非還元末端側から切断するセロビオヒドロラーゼ(EC 3.2.1.176)と二つのセロビオヒドロラーゼに分けて書きましょう。
1. Igarashi, et al., Science 333: 1279(2011), JBC 284: 36186 (2009), Methods Enzymol. 510: 169 (2012)
GH48の立体構造、ちょうど中央にトンネルが見え、ここへ基質であるセルロース鎖が入る。(Clostridium cellulolyticum 由来、PDB 1FAE、RasMolを使用して作成)
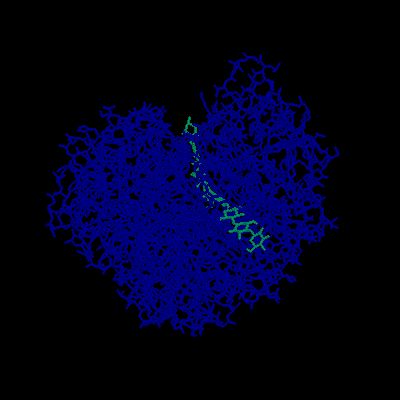
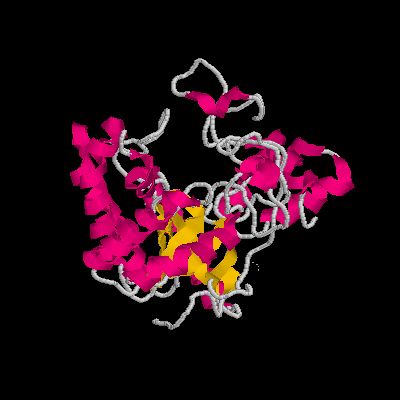
GH6の立体構造、やはり中央にトンネル構造があり、ここへ基質が入る。(Trichoderma reesei 由来セロビオヒドロラーゼII PDB 1OC7)